International Journal of Environmental Pollution and Remediation (IJEPR)
ISSN: 1929-2732

Volume 2 - Year 2014 - Pages 30-36
DOI: 10.11159/ijepr.2014.005
Phorbol Esters Degradation and Enzyme Production by Bacillus using Jatropha Seed Cake as Substrate
Chin-Feng Chang1, Jen-Hsien Weng1, Kao-Yung Lin2, Li-Yun Liu3, Shang-Shyng Yang4*
1Department of Food Science, China University of Science and Technology,
Nankang, Taipei 11581, Taiwan
afengb_b@yahoo.com.tw; toshibasl650@gmail.com
2Department of Living Science, National Open University,
Luzhou, New Taipei City 24701, Taiwan
lgu@mail.nou.edu.tw
3Department of Food Science, Nutrition and Nutraceutical Biotechnology,
Shih Chien University, Taipei 10464, Taiwan
liyun@mail.usc.edu.tw
4Department of Food Science, China University of Science and Technology,
Nankang, Taipei 11581, Taiwan, & Department of Biochemical Science and Technology, National Taiwan University,
Taipei 10617, Taiwan
ssyang@ntu.edu.tw
Abstract - The purposes of this research were to evaluate phorbol esters (PEs) degradation rate and enzyme production yield using submerged fermentation (SMF) as screening method and further using solid-state fermentation (SSF) as pilot scale-up study. SMF was carried out with 20 g seed cake in 100 ml minimal salt medium for 7 days incubation, while SSF was done with 20 g seed cake at 50% moisture content for 9 days incubation. Bacillus strains grew well on J. curcas seed cake with 108-1011 CFU/ ml in SMF for 3 days incubation, while they were 108-1010 CFU/ g in SSF. PEs reduced 76.5%, 77.1%, 78.4%, 85.5%, and 92.0% in SMF with B. smithii G16, B. sonorensis D12, B. licheniformis A3, B. subtilis H8 and B. coagulans C45 for 3 days incubation, respectively, and PEs completed degraded by these five strains for 7 days incubation. Maximum amylase, cellulase, lipase, pectinase, protease and xylanase productions in SMF were observed in B. sonorensis D12 (5.49 ± 0.49 U/ ml; day 7), B. subtilis H8 (17.03 ± 4.90 U/ ml; day 2), B. licheniformis A3 (59.03 ± 0.26 U/ ml; day 7), B. sonorensis D12 (1.70 ± 0.04 U/ ml; day 3), B. coagulans C45 (15.95 ± 0.35 U/ ml; day 7) and B. smithii G16 (1.40 ± 0.01 U/ ml; day 3), respectively. For SSF, PEs were reduced 86.0%, 83.2%, and 93.0% with B. sonorensis D12, B. subtilis H8 and B. smithii G16 for 3 days incubation, respectively. Maximum amylase, cellulase, lipase, pectinase, protease and xylanase productions in SSF were observed in B. smithii G16 (16.08 ± 0.36 U/ g; day 4), B. sonorensis D12 (2.94 ± 0.06 U/ g; day 2), B. smithii G16 (3.87 ± 0.64 U/ g; day 4), B. sonorensis D12 (8.13 ± 1.06 U/ g; day 2), B. smithii G16 (14.13 ± 0.30 U/ g; day 4) and B. smithii G16 (9.72 ± 0.97 U/ g; day 3), respectively. J. curcas seed cake could be detoxified by Bacillus and the high-protein seed cake could be potentially used for enzyme production in industry.
Keywords: Jatropha curcas seed cake, phorbol esters, degradation, submerged and solid state fermentation, enzyme production.
© Copyright 2014 Authors - This is an Open Access article published under the Creative Commons Attribution License terms. Unrestricted use, distribution, and reproduction in any medium are permitted, provided the original work is properly cited.
Date Received: 2013-09-12
Date Accepted: 2014-02-23
Date Published: 2014-03-20
1. Introduction
Due to the demand of local energy production, Jatropha curcas has high adaption capacity for land fertilization and can grow in marginalized land. It does not competitive with other vegetation crops for agricultural lands. The oil of J. curcas seed can be used as the raw materials for biodiesel production. However, biodiesel production from J. curcas seeds generate large quantum of residual de-oiled seed cake with an average of 700 g cake per kg of seed used [23]. The de-oiled J. curcas seed cakes contain high protein and other different nutrients like minerals, and amino acids etc. [1]. They can be used as the nutrient for animal feeds or production of valuable products. However, the de-oiled J. curcas seed cake contains anti-nutritional substances ex. trypsin inhibitors, curcins, tannins, saponins, phytates and toxic factors - phorbol esters (PEs) that restricts the uses of the seed cake. PEs have been identified as the main toxicants in cake which cannot be destroyed even by heating at 160oC for 30 min [23].
Physical methods, chemical treatments and microbial fermentation have been used to detoxify the toxic compounds in the seed cake ([4], [12], [17], [20], [22], [29]). However, the biological method would be more advantageous than the others for environmental friendly with safety and energy concerns. Especially solid state fermentation (SSF) could detoxify PEs and use for production of valuable products [17].
SSF and SMF techniques are common and conventional biotechnology processes for production of value-added products such as enzymes, biopharmaceuticals, organic acids, biosurfactants, vitamins, flavoring compounds, biofuels, biopesticides etc. [10]. SSF has economical and practical advantages over SMF, which includes use of raw materials as substrates, low capital cost, low energy expenditure, and less expensive downstream processing [6]. Therefore, SSF is adapted as commercial production and SMF is adapted as screening method for basic research.
In comparison with the previous works, Bacillus species were often subjected to the enzyme production (30) and most of Bacillus species are considered as Generally Regarded as Safe (GRAS) by FDA of USA. Additionally, Bacillus spp. was often using in production of food additives and probiotic products ([14], [32]). Therefore, Bacillus strains were applied to evaluate the degradation efficiency of PEs toxins factors in J. curcas seed cake, and J. curcas seed cake was used as the substrate for enzyme production in this paper.
2. Materials and Methods
2. 1. J. curcas Seed Cake and Tested Microbes
De-oiled J. curcas seed cake was kindly provided by Shin-Feng Energy Technology Co., LTD (Pingtung, Taiwan), then ground and pressed the defatted J. curcas seed cake. Five Bacillus spp. (B. coagulans C45, B. licheniformis A3, B. smithii G16, B. sonorensis D12, and B. subtilis H8) isolated from different compost plants and biofertilizers were used for PEs degradation and enzyme production [7].
2. 2. Biodegradation of PEs and Enzyme Production of J. curcas Seed Cake by Bacillus
For SMF, one ml of inoculums (about 108 CFU/ ml) was inoculated to 100 ml of mineral salts medium (MSM). Each litter of MSM medium contained seed cake 200 g, Na2HPO4.7H2O 6.7 g, KH2PO4 1.5 g, (NH4)2SO4 1 g, MgSO4.7H2O 0.2 g, ferrous ammonium citrate 0.06 g, CaCl2.2H2O 0.01 g and trace-element solution 1 ml. The trace element solution contained H3BO3 0.3 g, CoCl2.6H2O 0.2 g, ZnSO4.7H2O 0.1 g, MnCl2.4H2O 0.03 g, NaMoO4.2H2O 0.03 g, NiCl2.6H2O 0.02 g and CuSO4.5H2O 0.01 g in 1 1itter of 0.1N HCl [28]. The cultures were incubated in rotary shaker at 25oC, 150 rpm for 0, 1, 2, 3 and 7 days. PEs degradation and enzyme production were determined.
For SSF, 1ml of freshly prepared bacterial cells (about 109 CFU/ ml) was transferred to each of 250-ml Erlenmeyer flasks containing 20 g sterile seed cake at 50% initial moisture content. The flasks were then manually shaken well and incubated at 25oC for 0, 1, 2, 3, 5, 7 and 9 days. PEs degradation and enzyme production were determined.
2. 3. Phorbol Ester Extraction and Analysis
PEs were extracted and determined by the modified method of Hass and Mittelbach [13]. The mixture of J. curcas crushed seed cake 5 g and 95% ethanol 20 ml was shaken at 200 rpm for 5 min, then centrifuged at 14,000 g for 5 min. The residue was extracted two additional times with 95% ethanol. The extract fractions were combined and dried under vacuum at 50oC. The dried extract was dissolved in 1 ml 95% ethanol and passed through a 0.2-μm membrane filter. Phorbol esters were analyzed by HPLC system (Thermo Separation Products, U.S.) consisted of an AS1000 autosampler, P2000 pump and UV1000 detector. The solvents were water and acetonitrile: start with 60% water and 40% acetonitrile for 15 min, then 25% water and 75% acetonitrile for the next 20 min, and finally 100% acetonitrile for the next 20 min. Separation was performed at room temperature (25oC) with flow rate 1.3 ml/ min. The detector wavelength was set at 280 nm. The results were expressed as equivalent to phorbol-12-myristate-13-acetate (PMA) (Sigma, U.K.) used as an external standard.
2. 4. Enzyme Extraction and Assay
For enzyme assay in SMF, the sample was centrifuged at 12,000 g for 30 min and the supernatant was used for enzyme activities determination. While in SSF, 5 g fermented substrate was extracted with 20 ml 0.1 M Tris-HCl buffer pH 8.0 at 200 rpm orbital shaking for 30 min. The suspension was then centrifuged at 12,000 g for 30 min and the supernatant was used for enzyme assay.
Amylase, cellulase, lipase, pectinase, protease, and xylanase activities were determined by the methods of Bernfeld [5], Hu et al. [15], Kilcawley et al. [19], Janani et al. [16], Shimogaki et al. (31) and Joshi and Khare [18], respectively.
2.5. Chemical Analysis
Moisture, pH, protein, fat, and ash contents were analyzed by the standard methods of Association of Official Analytical Chemists [3]. Total organic carbon (TOC) was determined by TOC-5000A total organic carbon analyzer (Shimadzu, Japan).
3. Results and Discussion
3.1. Chemical Composition and Phorbol Ester Content in J. curcas Seed Cake
The chemical compositions of J. curcas seed cakes from Pingtung County, Taiwan are shown in Table 1. J. curcas seed cakes had pH 6.58 ± 0.02, moisture 5.92 ± 0.10%, ash 6.15 ± 0.20%, protein 24.24 ± 2.13%, fat 7.72 ± 0.26%, TOC 44.41 ± 0.01%, C/N ratio 11.02 ± 1.23 and PEs 0.60 ± 0.06 mg/ g dry sample.
PEs concentrations in seed cakes were lower than those of unshelled seed cakes from Nicaragua (1.78 mg/ g dry sample) [2], and Zimbabwe (0.70 mg/ g dry sample) [8], but they were higher than seed cakes from four provinces of Thailand (0.21-0.47 mg/ g dry sample) [11] This is possibly caused by various amounts of residual oil left in the samples and the variations of J. curcas in cultivation areas, soils, and climatic conditions ([4], [23], [26]).
Table 1. Chemical composition of J. curcas seed cakes.
|
Composition |
Amount |
|
pH |
6.58 ± 0.02 |
|
Moisture (%) |
5.92 ± 0.10 |
|
Ash (%) |
6.15 ± 0.20 |
|
Protein (%) |
24.24 ± 2.13 |
|
Fat (%) |
7.72 ± 0.26 |
|
TOC (%) |
44.41 ± 0.01 |
|
C/N ratio |
11.02 ± 1.23 |
|
Phorbol ester (mg g-1 dry sample) |
0.60 ± 0.06 |
|
|
Means ± S. D. (n=3) |
3.2. Degradation of Phorbol Esters and Enzyme Production by Submerged Fermentation
The J. curcas seed cake 20 g in 100 ml MSM medium was fermented with 5 Bacillus strains at 25oC. The Bacillus strains had initial cell numbers 104-106 CFU/ ml and then increased to 108-1011 CFU/ ml after 7 days incubation. While the PEs contents reduced 76.5%, 77.1%, 78.4%, 85.5%, and 92.0% with B. smithii G16, B. sonorensis D12, B. licheniformis A3, B. subtilis H8, and B. coagulans C45 for 3 days incubation, respectively (Figure 1).
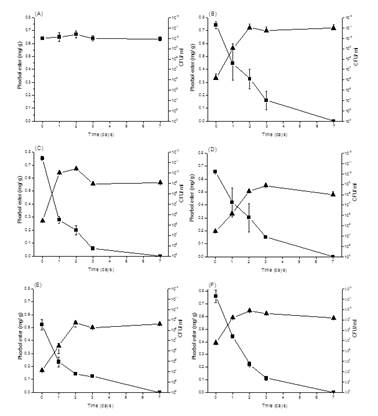
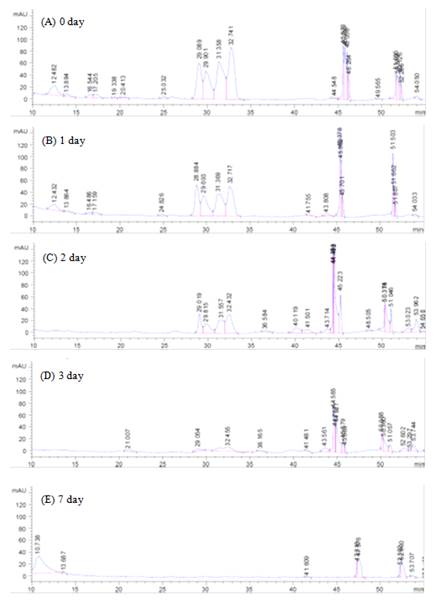
PEs contents in deoiled J. curcas seed cake were extracted by ethanol and analyzed by HPLC. The chromatogram (Figure 2) shows four major peaks of phorbol esters at retention times of 29.09-32.74 min, closely related to the reported by Makkar et al. [23]. PEs could be completely degraded by these five strains for seven days cultivation (Figures 1 and 2).
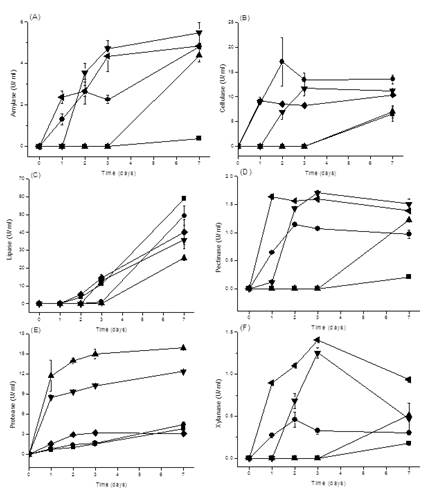
Enzyme productions of Bacillus strains in SMF at 25oC were shown in Figure 3. B. sonorensis D12 had the maximum amylase production 5.49 ± 0.49 U/ ml for 7 days cultivation. B. subtilis H8 had the maximum cellulase production 17.03 ± 4.90 U/ ml for 2 days cultivation. B. licheniformis A3 had the maximum lipase production 59.03 ± 0.26 U/ ml for 7 days cultivation. B. sonorensis D12 had the maximum pectinase production 1.70 ± 0.04 U/ ml for 3 days cultivation. B. coagulans C45 had the maximum protease production 15.95 ± 0.35 U/ ml for 7 days cultivation. B. smithii G16 had the maximum xylanase production 1.40 ± 0.0 U/ ml for 3 days cultivation.
3.3 Degradation of Phorbol Esters and Enzyme Production by Solid-state Fermentation
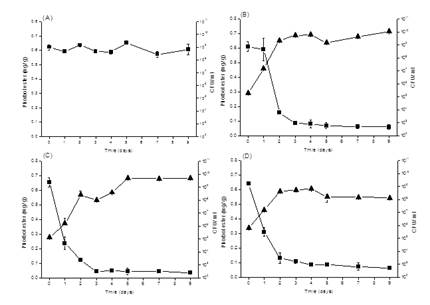
For SSF, 20 g seed cake with 50% initial moisture content at 25oC was fermented with 3 Bacillus spp. (H8, D12 and G16) had high enzyme production in SMF. The Bacillus strains had initial cell numbers 104-106 CFU/ g and then increased to 108-1011 CFU/ g after 7 days incubation. The PEs contents in seed cake reduced 86.0%, 83.2% and 93.0% with B. sonorensis D12, B. subtilis H8, and B. smithii G16 for 3 days incubation at 25oC, respectively (Figure 4).
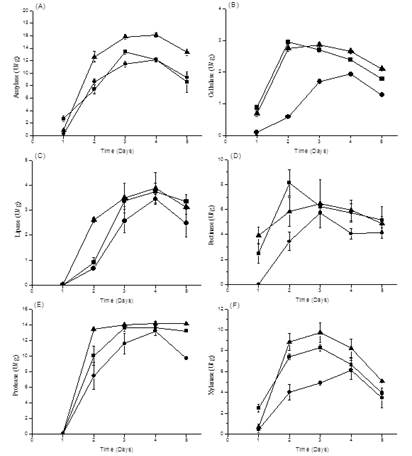
The enzyme productions of tested microbes increased with cultivation and had the maximal production for 2-4 days cultivation (Figure 5). B. smithii G16 had the maximum amylase production 16.08 ± 0.36 U/ g for 4 days cultivation. B. sonorensis D12 had the maximum cellulase production 2.94 ± 0.06 U/ g for 2 days cultivation. B. smithii G16 had the maximum lipase production 3.87 ± 0.64 U/ g for 4 days cultivation. B. sonorensis D12 had the maximum pectinase production 8.13 ± 1.06 U/ g for 2 days cultivation. B. smithii G16 had the maximum protease production 14.13 ± 0.30 U/ g for 4 days cultivation. B. smithii G16 had the maximum xylanase production 9.72 ± 0.97 U/ g for 3 days cultivation.
The world market of industrial enzymes is estimated to be 1.6 billion $US, including food enzymes (29%), feed enzymes (15%), and general technical enzymes (56%). Bacillus can secrete a variety of soluble extracellular enzymes [27] and most of Bacillus species are considered as GRAS. It is estimated that Bacillus spp. enzymes make up about 50% of the total enzyme market around the world [25]. Therefore, Bacillus is a good candidate microbe for enzyme production of J. curcas seed cake. Each Bacillus strain in this study has different capacities of enzyme production, and can be applied by SMF or SSF for enzyme production in the future.
Phengnuam and Suntornsuk [26] reported that B. licheniformis and B. subtilis could degrade PEs 60% and 40% for 5 days incubation with SMF. In this study, 5 Bacillus strains can degradation PEs 76.5-92.0% for 3 days incubation and 100% for 7 days incubation by SMF. Joshi et al. [17] reported that Pseudomonas aeruginosa degradated 60-73% phorbol esters in the seed cake for 6 days incubation with SSF, and Belewu and Sam [4] also reported that Aspergillus niger degraded 77% phorbol esters in the seed cake for 7 days incubation by SSF. In this study, 3 Bacillus strains can degrade PEs 83.2-93.0% for 3 days incubation by SSF. Both of SMF and SSF can be used for PEs degradation [26].
Jatropha seed oil has been used extensively for bio-diesel production. The residue of J. curcas seed cake can be used as substrate in SMF or SSF for production of useful substances. In SMF, Choudhury et al. [9] used J. curcas seed cake as nutrients for pullulan production by an osmotolerant yeast Aureobasidium pullulans. Phengnuam and Suntornsuk [26] used J. curcas seed cake as nutrients for protease, phytase, and esterase production by Bacillus licheniformis. In SSF, different kinds of microbes like Aspergillus niger, Aspergillus oryzae, Pseudomonas aeruginosa, and Scytalidium thermophilum were used for xylanase, cellulase, protease, and lipase production ([18], [21], [24], [33]). From this study, it showed that SMF and SSF could degrade PEs efficiency, and the high-protein seed cake could be used for enzyme production or other valuable products production in the future.
References
[1] P.C. Abhilash, P. Srivasatav, S. Jamil "Revisited Jatropha curcas as an oil plant of multiple benefits: critical research needs and prospects for the future" Environmental Science and Pollution Research, 18, 2011, 127-131. View Article
[2] E.M. Aregheore, K. Becker, H.P.S. Makkar "Detoxification of a toxic variety of Jatropha curcas using heat and chemical treatments, and preliminary nutritional evaluation with rats" The South Pacific Journal of Natural Science, 21, 2003, 50-56. View Article
[3] Association of Official Analytical Chemists (AOAC) "Official Methods of Analysis of AOAC International, 16th edition. AOAC International" 1998, Gaithersburg, MD, USA.
[4] M.A. Belewu, R. Sam "Solid state fermentation of Jatropha curcas kernel cake: proximate composition and antinutritional components" Journal of Yeast and Fungal Research, 1, 2010, 44-46. View Article
[5] P. Bernfeld "Amylase α & and β. In Methods in Enzymology" Academic Press Inc., New York, 1955, pp. 149-158. View Article
[6] L.R. Castilho, C.M.S. Polato, E.A. Baruque, G.L. Sant'Anna Jr, D.M.G. Freire "Economic analysis of lipase production by Penicillium restrictum in solid-state and submerged fermentations" Biochemical Engineering Journal, 4, 2000, 239-247. View Article
[7] C.H. Chang, S.S. Yang "Thermo-tolerant phosphate-solubilizing microbes for multi-functional biofertilizer preparation" Bioresource Technology Journal, 100, 2009, 1648-1658. View Article
[8] E. Chivandi, J.P. Mtimuni, J.S. Read, S.M. Makuza "Effect of processing method on phorbol esters concentration, total phenolics, trypsin inhibitor activity and the proximate composition of the Zimbabwean Jatropha curcas provenance: A potential livestock feed" Pakistan Journal of Biological Sciences, 7, 2004, 1001-1005. View Article
[9] A.R. Choudhury, N. Sharma, G.S. Prasad "Deoiled jatropha seed cake is a useful nutrient for pullulan production" Microbial Cell Factories, 11, 2012, 39-47. View Article
[10] S.R. Couto, M.A. Sanromán "Application of solid-state fermentation to ligninolytic enzyme production" Biochemical Engineering Journal, 22, 2005, 211-219. View Article
[11] S. Donlaporn, W. Suntornsuk "Toxic compound, anti-nutritional factors and functional properties of protein isolated from detoxified Jatropha curcas seed cake" International Journal of Molecular Science, 12, 2011, 66-77. View Article
[12] G. Goel, H.P.S. Makkar, G. Francis, K. Becker "Phorbol esters: structure, biological activity, and toxicity in animals" International Journal of Toxicology, 26, 2007, 279-288. View Article
[13] W. Hass, M. Mittelbach "Detoxification experiments with the seed oil from Jatropha curcas L" Industrial Crops and Products,12, 2000, 111-118. View Article
[14] H.A. Hong, L.H. Duc, S.M. Cutting "The use of bacterial spore formers as probiotics" FEMS Microbiology Reviews, 29, 2005, 813-835. View Article
[15] C.C. Hu, L.Y. Liu, S.S. Yang "Protein enrichment, cellulase production and in vitro digestion improvement of pangolagrass with solid state fermentation" Journal of Microbiology, Immunology, and Infection, 45, 2012, 7-14. View Article
[16] L.K. Janani, G. Kumar, K.V. Bhaskara Rao "Screening of pectinase producing microorganisms from agricultural waste dump soil" Asian Journal of Biochemical and Pharmaceutical Research, 1, 2011, 329-336. View Article
[17] C. Joshi, P. Mathur, S.K. Khare "Degradation of phorbol esters by Pseudomonas aeruginosa PseA during solid-state fermentation of deoiled Jatropha curcas seed cake" Bioresource Technology, 102, 2011, 4815-4819. View Article
[18] C. Joshi, S.K. Khare "Utilization of deoiled Jatropha curcas seed cake for production of xylanase from thermophilic Scytalidium thermophilum" Bioresource Technology, 102, 2011, 1722-1726. View Article
[19] K.N. Kilcawley, M.G. Wilkinson, P.F. Fox "Determination of key enzyme activities in commercial peptidase and lipase preparations from microbial or animal sources" Enzyme and Microbial Technology, 31, 2002, 310-320. View Article
[20] A. Kumar, S. Sharma "An evaluation of multipurpose oil seed crop for industrial uses (Jatropha curcas L.): a review" Industrial Crops and Products, 28, 2008, 1-10. View Article
[21] N. Mahanta, A. Gupta, S.K. Khare "Production of protease and lipase by solvent tolerant Pseudomonas aeruginosa PseA in solid-state fermentation using Jatropha curcas seed cake as substrate" Bioresource Technology, 99, 2008, 1729-1735. View Article
[22] H.P.S. Makkar, A.O. Aderibigbe, K. Becker "Comparative evaluation of nontoxic and toxic varieties of Jatropha curcas for chemical composition, digestibility, protein degradability and toxic factors" Food Chemistry, 62, 1998, 207-215. View Article
[23] H.P.S. Makkar, K. Becker, F. Sporer, M. Wink "Studies on nutritive potential and toxic constituents of different provenances of Jatropha curcas" Journal of Agricultural and Food Chemistry, 45, 1997, 3152-3157. View Article
[24] T. Ncube, R.L. Howard, E.K. Abotsi, E.L.J. van Rensburg, I. Ncube "Jatropha curcas seed cake as substrate for production of xylanase and cellulase by Aspergillus niger FGSCA733 in solid-state fermentation" Industrial Crops and Products, 37, 2012, 118-123. View Article
[25] H. Outtrup, S.T. Jorgensen "The importance of Bacillus species in the production of industrial enzymes" In Applications and systems of Bacillus and relatives. Blackwell Science Inc., 2002, Malden, Mass. pp. 206-218. View Article
[26] T. Phengnuam, W. Suntornsuk "Detoxification and anti-nutrients reduction of Jatropha curcas seed cake by Bacillus fermentation" Journal of Bioscience and Bioengineering, 115, 2012, 168-172. View Article
[27] F.G. Priest "Extracellular enzyme synthesis in the genus Bacillus" Bacteriology Review, 41, 1977, 711-753. View Article
[28] B.A. Ramsay, K. Lomaliza, C. Chavarie, B. Dubé, P. Bataille, J.A. Ramsay "Production of poly-(Β-hydroxybutyric-co-Β-hydroxyvaleric) acids" Applied and Environmental Microbiology, 56, 1990, 2093-2098. View Article
[29] D. Saetae, W. Suntornsuk "Antifungal activities of ethanolic extraction from Jatropha curcas seed cake" Journal of Microbiology and Biotechnology, 20, 2010, 319-324. View Article
[30] M. Schallmey, A. Singh, O.P. Ward "Developments in the use of Bacillus species for industrial production" Canadian Journal of Microbiology, 50, 2004, 1-17. View Article
[31] H. Shimogaki, K. Takeuchi, T. Nishino, M. Ohdera, T. Kudo, K. Ohba, M. Iwama, M. Irie "Purification and properties of a novel surface active agent and alkaline-resistant protease from Bacillus sp. Y" Agricultural and Biological Chemistry, 55, 1991, 2251-2258. View Article
[32] M.L. Teixeira, F. Cladera-Olivera, J. dos Santos, A. Brandelli "Purification and characterization of a peptide from Bacillus licheniformis showing dual antimicrobial and emulsifying activities" Food Research International, 42, 2009, 63-68. View Article
[33] A. Thanapimmetha, A. Luadsongkrama, B. Titapiwatanakunc, P. Srinophakuna "Value added waste of Jatropha curcas residue: Optimization of protease production in solid state fermentation by Taguchi DOE methodology" Industrial Crops and Products, 37, 2012, 1-5. View Article